|
回主页
第七节 染色体数目的差异是猿进化成人的鸿沟
第6节中,我们已经得出了这样的结论:全人类源于最早的一对父母--科学亚当和科学夏娃。他们是由猿进化来的吗?在人类学的现代科学上是怎样看的呢?这就是本节要讨论的问题。
一.人类是由进化产生的论点
1. 建立在“群体的、逐渐的”进化论的基点
达尔文进化论的基点是“代代相傳,略有差異”(descent with modification)。意即每世代都会產生許多有变異的个体,他們的特征可以被下一代继承。并且,由于许多代变异的积累,形成了适应自然能力不同的群体。当自然环境变化时,一些不能适应的群体灭绝了,能适应的群体留下了,久而久之,形成了新的物种,这就是进化。
对于人来源于进化的描述大致是:第三纪的中新世开始变冷,森林范围缩小,部分被毁灭,出现草原和荒漠。原住在森林中的古人猿逃向赤道地区,住在森林边缘和平原上。于是“用四肢攀缘,林栖,主要吃果实的动物”,逐渐进化为能够两足直立行走、杂食、狩猎的动物。能够适
应环境变化的就保存下来,继续进化发展,不能适应的就灭绝了。第四纪更新世冰期来临,冰期和间冰期的气候冷暖和干、湿交替变化,植物覆盖、冰川、河流以及海、陆发生相应的变迁,大大促进了人类的进化。得以适应变化巨大的生存环境的,被留了下来,终于形成了人类。在人们不知道染色体和DNA时,这一切的描述显得十分合理。
2.无法逾越的鸿沟
进化论认为人与黑猩猩是共祖的。因为人进化了,而黑猩猩没有进化,所以在600多万年前分道扬镳。按照这个假定,这共同的祖先(古人猿)就应该是和黑猩猩(包括猩猩、大猩猩和红毛猩猩)一样,有24对染色体。有了高倍显微镜后,人们看到了细胞中的染色体,认识了不同的物种,往往有不同数量的染色体。
1923年,美国遗传学权威、得克萨斯大学校长西奥菲勒斯·佩音特(Theophilus Painter)提出人体的染色体数目为2n=48,这被后来
作为一条定论充斥于各种教科书和百科全书。直到1956年美籍华裔学者蒋有兴(Joe Tjio JH)、徐道觉(Hsu TC)和Albert Levan才首先正确鉴定了人类染色体是2n=46条而不是48条。
由此可以推断,24对染色体的古人猿与23对染色体的现代人之间有一个鸿沟,染色体对数的区别就阻断了古人猿进化成人的可能性。因为达尔文认为进化是“无数的、连续的、轻微的变异”(by
numerous, successive, slight modifications)渐变形成的,而染色体对数的差别,成为了靠着“渐变进化”无法跨越的鸿沟。就像不可能去商店购买47.99个鸡蛋,染色体条数同样不能以非整数存在,因此,不能有染色体条数的数目一点点地渐变。例如不可能在48条 (24对)染色体和46条(23对)染色体中间,存在着47.99、47.98、47.97… 等染色体条数的、渐变过程中的猿人。
达尔文在“物种起源”中说:“徜使能证明有任何复杂器官不是经过无数的、连续的、轻微的变异形成的,那么我的学说就要完全破产。但是,我还没有发见这种情形。”
中文版 第72页
人类23对染色体集合的细胞核,可以说是一种基本的、微小的“复杂器官”,也是所有器官构成的核心,恰恰它就是“不可能”从古人猿的24对染色体的细胞核“经由无数、连续地、微小的变异而形成”的。24对染色体的各种器官,也都不可能如此地“进化形成”23对染色体的器官,难道不是这样吗?达尔文没有能发现是十分合理的,因为他没有高倍显微镜。但是,达尔文的这句话,很客观地为他的理论留下了余地。今天,他留下的余地被应验了,结论是:他的“学说就要完全破产”了。
在同一书中还说:“因为自然选择只是利用微细的、连续的变异而发生作用;她从来不能采取巨大而突然的飞跃,而一定是以短的、确实的、虽然是缓慢的步骤前进。”中文版
第76页
那么,古人猿如何经由“微小而可靠的,虽然是缓慢的步骤”来跨越这个染色体数目差异的鸿沟呢?显然是不可能的。
3.从染色体、DNA看物种的隔离
仅仅是在近50年来,通过生物科学的研究,才对染色体和DNA有比较清晰的认识。我们知道,“科学是有关物质世界及其现象,并需要用到客观观察和系统实验的知识体系。”
(见《大英百科全书》
而在自然科学的知识体系方面,必须借助不同时期的技术手段来达到。一百多年前,在认定人是由古猿进化而来时,人们根本就没有高倍显微镜,也没有分析DNA的技术。可以看出,那时的认识,必然受着当时的科学技术手段所局限。
例如,马和驴交配可以产生出骡子,但公骡和母骡的交配不会产生小骡子,以前是认为“母骡后面盆骨不能开合,所以不能产子”,现在知道这是因为染色体数形成的物种隔离。马有64条染色体,驴有62条对染色体,它们交配生下的骡子是63条染色体。骡子没有生殖能力是因为染色体不成双(63个是单数),生殖细胞无法进行正常的分裂(减数分裂)。在非常罕见的情况下,骡子有可能和马(或驴)交配生出马驹(或驴驹),这新的一代染色体一定是64条或62条,它只是复原成了它的祖辈,但不可能形成一个新的物种。
其实人当中也有染色体畸变病的患者,他们只有45条(23.5对)染色体。这样的病人从外表看来,也有可能是正常的,但很难生育下一代,只有较小的机会与46条染色体的人生出正常的人,但不可能形成一个新的人种。
狮子和老虎都是38条(19对)染色体,它们互相交配有很小概率孕育生出“狮虎”(或“虎狮”)。“狮虎”也有更小的可能与狮子或老虎生出下一代,那将是一个正常的狮子或老虎,并没有发现过一个“狮虎”的新物种。为什么相同染色体数目的狮子和老虎不能交配产生一个新的物种呢?这深层的DNA原因还需要人们的继续探索,但是,由于DNA的隔离,不能产生出一个“狮虎”新物种却是一个不争的事实。
4.新进化论的理论也无法挽救进化论
现在出现了一个“综合进化理论”,试图用基因和DNA突变来詮解古老的进化论。然而,无论是突变、遗传漂变和基因流动等因素,都不可能实现古人猿经由“微小而可靠的,虽然是缓慢的步骤”,去改变染色体的对数。23对染色体和24对染色体对数差异,仍然是这些新进化论无法逾越的鸿沟。
二.骤变产生“23对染色体人的始祖”的假说
面对进化论无法解决的染色体鸿沟问题,有人重新提出了物种产生的“骤变论”,认为是古人猿的染色体发生了骤变,24对染色体瞬间变成了23对,产生了两个23对染色体的现代人始祖,并以此跨过了染色体对数的鸿沟。
骤变论(Saltation :Jumping and discontinuous changes)认为生物的变异,是“非渐进”的,甚至只需要一个步骤便能形成新物种。提出骤变论的重要人物之一是米瓦特(St. George Jackson Mivart
,1827–1900),他是一位和达尔文同时代的生物学家。米瓦特相信新物种“是突然的,而且是一蹴而成”产生的(“with
suddenness and by modifications appearing at once.”)。1871年,米瓦特出版了他的著作《在物种的成因》(On the Genesis
of Species)和达尔文进行了针锋相对的争论,我们在第九节还要详细叙述。
1.建立在DNA“骤变” 假说上的基点
这个假说是建立在以下三个基点上:
(1)人类和黑猩猩有共同祖先:24对染色体的古人猿。黑猩猩没有进化,仍保持了24对染色体;
(2)由于骤变,古人猿的24对染色体中的两个染色体发生了融合,变成了人的2号染色体,因此产生了具有23对染色体的人类;
(3)这种因骤变产生染色体融合的现象,可以参考的是在目前人类中仍然存在着的染色体病:罗伯逊易位病 ( Robertsonian
translocation)。
2. 罗伯逊易位病
根据着丝粒的位置,人类染色体可以分为三种:(1)近中着丝粒染色体,着丝粒位于或靠近染色体中央,将染色体分为长短相近的两个臂;(2)亚中着丝粒染色体,着丝粒偏于一端,将染色体分为长短明显不同的两个臂;(3)近端着丝粒染色体,着丝粒非常靠近染色体的一个顶端。
罗伯逊易位是一种常见的染色体疾病,其发病率约为千分之一,都是“近端着丝粒染色体”的融合。在人类细胞核中的23对染色体中,有5对“近端着丝粒染色体”,如图7-2中的13、14、 15、 21 和
22号染色体。
罗伯逊易位是在“近端着丝粒染色体”的“着丝粒处”或其附近断裂后,两个染色体融合为一个染色体,形成了异常的染色体结构。例如,在生殖细胞“减数分裂”产生精子时,其第13号染色体和第14号染色体的长臂融合成了一个染色体(它们的短臂丢失),那么这个精子中,只包含了22条染色体。因为“近端着丝粒染色体”的短臂相当短,丢失的遗传信息不多,这个精子仍然可以产生受孕。当这个精子和一个正常卵子(包含23条染色体)结合时,就会产生出45条染色体的胚胎。当这个胚胎发育成长成人后,仍然可以是一个基本正常(医学上叫表型正常--Phenotype
normal[注释1])的人,只是这样的病人即使和正常染色体的配偶结合,在孕育时,常常会发生自然流产、死胎、畸形生育等病症,只有很小的几率生出正常的、23对染色体的孩子。
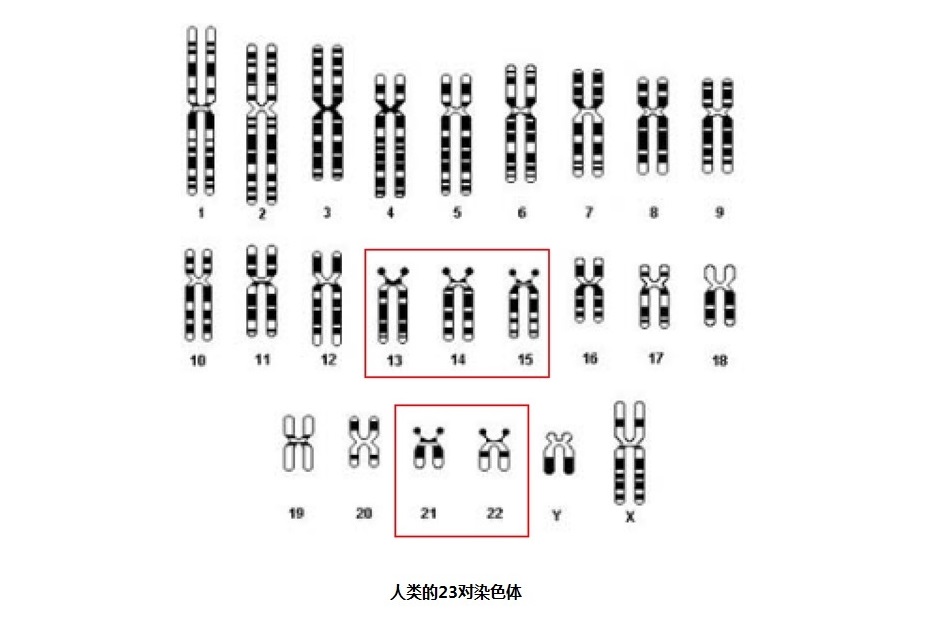
图7-1 人类的23对染色体的着丝粒位置图
3.人类的2号染色体
2005年4月,在英国《自然》杂志上,“美国国家人类基因组研究所”发布了由122位研究人员签名的研究报告“Generation
and annotation of the D NA sequences of human chromosomes 2 and 4”“人类2号和4号染色体全面注解的序列”,这是世界基因组研究计划中的一个项目。此报告应该是对2号染色体最权威的研究成果 。
在报告的结论中有这样的说明:“在2号染色体上,过去曾经描述过,在2q13–2q14.1位置上,有围绕着祖先染色体融合的区域。在这里,我们鉴定出2q21.1-2q21.2内一段2.6-Mb区域内有着丰富的近着丝粒、重复和着丝粒卫星重复序列”,这“也许是标志着祖先的端粒和着丝粒,可能代表着人类世系中的常染色体的经常性重组。”
“On chromosome 2, the local region surrounding the ancestral
chromosomal Fusion site on 2q13–2q14.1 had previously been described. Here we
identified a 2.6-Mb region within 2q21.1–2q21.2
that is enriched for pericentromeric duplications
and centromeric satellite repeat sequence motifs”,“ perhaps representative of
recurrent chromosome rearrangements in the human lineage.”
在第6节中,我们已经质疑了“在2q13–2q14.1位置上”所谓的融合区的“TTAGGG”序列,知道它不可能是表达两个染色体端粒的融合。但是在这里,我们仍然按照骤变论的设想,继续的来进行分析。
根据这个报告结论,假定了人的2号染色体,是由两个古人猿的染色体融合而成。参照罗伯逊易位的病理和上面结论中所描述的融合点的位置、退化着丝粒的位置,研究人员绘制了图片(图7-2),表达了它们的融合的方式。
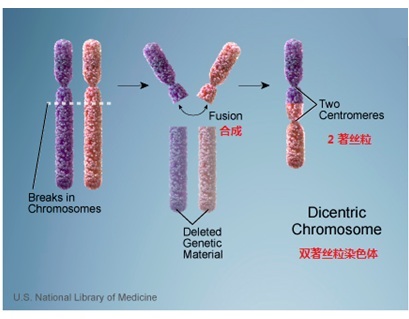
图7-2 骤变论对2号染色体合成的假想图
按照上面报告的结论和此图,可以在人类2号染色体的结构图上,明确地划分出融合点和残留着丝粒的位置(图7-3)。
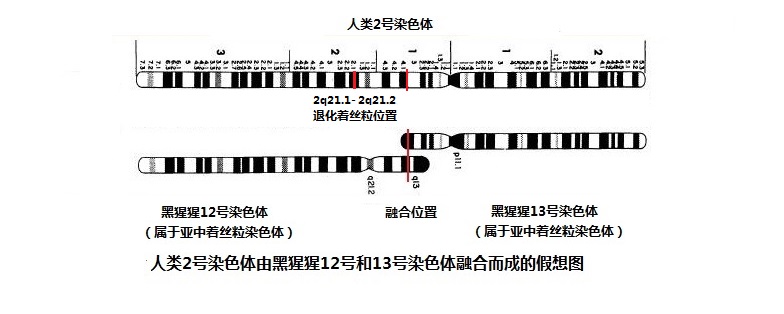
图7-3 2号染色体融合假想图(这只是一个假想图,您不能把它看成真的)
|