|
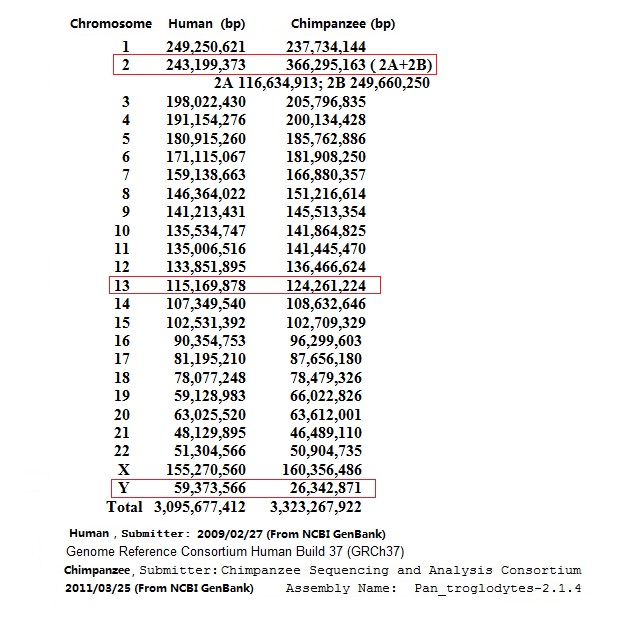
4.�ں����۽�����һ������ ��1���˺ͺ�����Ⱦɫ��IJ�� ͼ7-4 �µĺ����ɵ�Ⱦɫ�����ݺ��˵�Ⱦɫ�����ݱȽ�������GenBank�� ͼ7-4 �У���ʾ������ͺ���������Ļ��������ݣ����������ݶ�����������NCBI
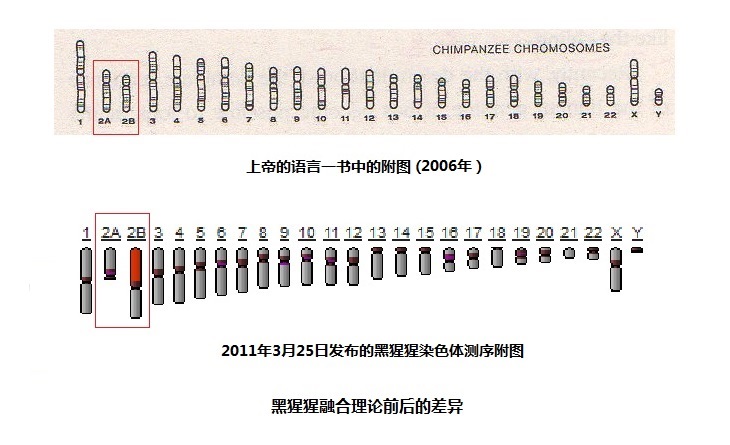
GenBank�� (��վ��http://www.ncbi.nlm.nih.gov/genome/202) ������Ի�����Ķ��廹������ô���������һ����õĴ𰸡���������ǰ���ȫ��Ⱦɫ���DNA�����������ݵļ��ϣ�����Ļ����������22�Գ�Ⱦɫ���һ����Ⱦɫ�壨XȾɫ���YȾɫ�壩���ܹ���3,095,677,412��������������ɵĻ����������23�Գ�Ⱦɫ������ǵ�һ����Ⱦɫ�壬�ܹ���3,323,267,922������ԡ��������ǿ������й��˺ͺ����ɻ�������ȷ�����ϡ� ͼ�к����ɻ��������ϣ����ɡ������ɲ�����������ˡ���2011��3�·����ģ������Ѿ���Ⱦɫ��ij��Ȳ��Ե���λ�������������Ϻ���ǰ��������ȣ������ɵĻ����������Ļ�����IJ��죬�ȹ�ȥ֪���ĸ��������ܳߴ�IJ�����1:1.07��YȾɫ��IJ��������1:0.44��13�ų�Ⱦɫ��IJ�����1:1.08�����ﷱ���Ŵ��У�DNA���ж������ϸ�ĸ��������ݡ�������DNA�������ݵIJ��죬�Լ�YȾɫ��������ȶ�����ױ仯��Ⱦɫ�壬����������IJ�𣬳��ط����˺ͺ����ɹ���Ŀ��ܡ� ��2������������Ⱦɫ���ںϵĿ��� ͼ7-5 �������ں�����ǰ��IJ��� �����2��Ⱦɫ���ɺ����ɵ�2A��2BȾɫ��ϳɵ���һ���룬����������������Ϊ�ںϵĸ��ݡ������ɵ�Ⱦɫ���ţ�Ӧ�ú���������һ��������Ⱦɫ��ij��������������һ����Ӧ����1�ţ��Ժ��ȱ�ŵݼ���Ϊʲô����Ϊ2A��2B�أ���Ϊ������Ϊ����������Ⱦɫ�壬�ںϳ������2��Ⱦɫ�塣 Ȼ�����ܶ�������������ˣ�������������ȷ�������������ɵ�Ⱦɫ����2A��2B��2011����ǰ������ȷ����2A��2B������������Ӧ���Ǻ����ɵ�12��Ⱦɫ���13��Ⱦɫ�壨ȥ��2A��2B��ͼ�е�3��Ⱦɫ��Ӧ����2��Ⱦɫ�壩������֮���䣬˵������Ⱦɫ��ij��ȼ�����������2��Ⱦɫ�峤�����������Ⱦɫ����ͷ��ͷ���ںϡ�����˵��������Ļ����Ǻ�����2��Ⱦɫ���ϵĻ������зdz���Ӧ���ڡ��ϵ۵����ԡ�һ���У����Բ�ͼ˵���� 2011�꣬�������ɲ�����������ˡ������IJ���ͼ������������ȫ��ͬ������Ⱦɫ���Ϊ��2A��2B���ӳ����Ͽ���2B��249,660,250������ԣ����Ӧ����1�ų�Ⱦɫ�壨�ο�ͼ7-4����2A��116,634,913������ԣ����Ӧ����14�ų�Ⱦɫ�塣����ǰ��������ļ�������̫���ˣ�����ô�������أ���������¼��룬�����ɵ�2A��2B����Ⱦɫ��Ҫ�ںϳ������2��Ⱦɫ�壬��Ҫ����3��֮һ��Ⱦɫ�峤�ȡ����ִ�ҽѧ�������������������IJ�̬�����ɸ��������ܴ����ڲ�����ȫ��ͬȾɫ����ϵ�һ�к�һŮ�������ˣ����Dz����ܵġ� ��Ŀǰ����ֹ���ڣ�2014��5��1�գ�����������2��Ⱦɫ��ͺ�����2A��2BȾɫ��ͼ�Ͽ�����2��Ⱦɫ��Ļ���������2787����������2A�Ļ��������������999����2B��994����������1993����������2��Ⱦɫ������Ļ�����Ŀ���Ⱥ�����2A��2BȾɫ��Ļ���������Ҫ���794������������ɵ�����Ⱦɫ���ں�ʱ��Ҫ��ʧ����֮һ������2��Ⱦɫ�������Ŀ�������ĸ��ࡣ��ô�������2��Ⱦɫ��������ǧ�����������������أ���ô���ھ��Ӻ������У�Ⱦɫ���ںϵ�һ˲�䣬������ô��Ļ�����˹�����������Ľ�����ר���ǣ�˭�ܽ����أ����ο������ĸ�¼���֣� �ɴ˿ɼ�����һ���ںϵļ��룬��ȫû���κο�ѧ���ݡ� 5������ѧ�ǵ����͡���ѧ���ޡ����ɸ��ʼ��� ��� ����ѧ�ǵ����͡���ѧ���ޡ�
���������ɹ���Գ�����������ƶ�����䡱����ѷ��λ���ֽ�������������IJ��䣬��ô���Dz����ĸ����ж���أ� ���ڲ����ܵõ�����Գ�����ϣ��ڹ���Ⱦɫ���ںϷ����������������ϣ����Ʋ����ִ��˵��й����ݡ��ڹ���Գ��ֳ����ʱ�κ����������ϣ��ο��ִ������ɵ����ϡ� �ִ�������5����������ѷ��λ����������˿��Ⱦɫ�塱�����2��Ⱦɫ����������������˿��Ⱦɫ�塱�ںϵļٶ�����ô������Գ��24��Ⱦɫ���У�Ӧ����7���ǡ�������˿��Ⱦɫ�塱����������������Ǻ����ɵ�12�ź�13��������Ϊ2A��2B����Ⱦɫ�塣
������5����������˿��Ⱦɫ�塱�������Ŵ�ѧ���ۣ��ɷ���15�ֲ�ͬ��ϵ���ѷ��λȾɫ�塣������ѷ��λ�����ı�����ǧ��֮һ����ô��ÿһ�ַ����ĸ��ʻ��С������Գ��7�ԡ�������˿��Ⱦɫ�塱�����пɷ���28�ֲ�ͬ��ϵ���ѷ��λȾɫ�塣��Ϊ�������ر����λ��Ⱦɫ���в����ѣ���������˿���У��ֱ�����һ��ʧȥ��˿�����ܣ�������⣬������ѷ��λ�ĸ��ʽ���С�����⣬��ͼ7-3�п��Կ���������Գ��������ɣ���12�ź�13��Ⱦɫ�岢������������˿��Ⱦɫ�塱�����ǡ�������˿��Ⱦɫ�塱������Ⱦɫ����λ������Ƶ��Զ���ڡ�������˿��Ⱦɫ�塱������Ƶ�ʡ�������ĵ�Ⱦɫ�岡�У����ٷ�������������˿��Ⱦɫ�塱���ںϡ�����Щ�������������Գ12�ź�13��Ⱦɫ����ںϷ����ʻ�ԶС��ǧ��֮һ�� ��������������ִ��˵���ѷ��λ��Ϊǰ��ģ�Ҳֻ�����ִ�����ѷ��λ������״����˼�����ٶ�����Գ�ںϳ�2��Ⱦɫ�����һ�������ƣ�����������������λ�������ʣ�Ϊȫ����ѷ��λ���������е����֮һ����ô���ڹ���Գ��ֳ�У�Լ����ǧ��֮һ��Ӥ�������ںϵ�2��Ⱦɫ����Ҳ����˵ÿ����5000��Ӥ���У���һ��47��Ⱦɫ��Ӥ��������������23��Ⱦɫ��ľ��ӣ�����һ��Ⱦɫ�壩��������24��Ⱦɫ������ӽ���������ĺ��ӡ�������23��Ⱦɫ������ӣ�����һ��Ⱦɫ�壩��������24��Ⱦɫ��ľ��ӽ���������ĺ��ӡ�����Ӥ����ϸ�����а�������47��Ⱦɫ�壨23��24�������Ǿ���23.5��Ⱦɫ�������Գ���ӡ�������Գ�˵ĺ��Ӿ������ڻ�����ѷ��λ�IJ���һ����������47����23.5�ԣ�Ⱦɫ�����һ������˲����д��ڽӴ��������� ���Ҫ����һ���ִ���������46��Ⱦɫ���Ӥ����������һ��23��Ⱦɫ��ľ���ǡ��������һ��23��Ⱦɫ������ӣ��������Ƕ�����Ϊ2��Ⱦɫ���ںϣ������ںϳ�������ŵ�Ⱦɫ�壩�����Բ�����һ��Ⱦɫ�塣��ˣ���������һ���ִ���������23��Ⱦɫ����ˣ�������������Գ���ӡ����������Ӧ����5ǧ��֮һ����5ǧ��֮һ�������2ǧ5�����֮һ��Ҳ����˵ÿ����2ǧ5���������ԳӤ�������л����һ��23��Ⱦɫ��ĵ�������Ů�ˡ� �ִ�������Ⱥ�����ԼΪ2ǧ����������һ�������У���Ȼʳ��Ĺ����������ƣ��������и����Ⱥ�塣��ͬ�������������ƹ���ԳȺ��������С���ֹ۵�ȷ��Ϊ1�������ÿ��ij��������������Ⱥ��������5%���㣨�й��ִ����ڼƻ�������ǰ�ij�����Ϊ2%��4.3%��ƽ��3.7%������ô���������ԳȺÿ���ܹ�����500��Ӥ����������ı�������ÿ����2ǧ5���������ԳӤ���������һ��23��Ⱦɫ��ĵ�������Ů�ˡ����ɴ����ۣ��˹���ԳȺÿ5���꣨2ǧ5�������500�����Բ���һ���ִ���������������Ů�ˡ��Ӹ����ϼ��㣬Ҫ����һ�����˺�һ��һŮ�ˣ�����Ҫ������5���꣬��10�����ʱ�䡣 ��һ����ѵ��ǣ����DZ���������һ���е����25�����������ʱ�����������మ��������������һ��һŮ���ο��������ɵ�����Լ40�꣩��������������Ŀ��ܡ����ǣ������Ƿֱ�����10��������ʱ��ij����У�����ÿ��ֻӵ���൱��һ˲���ŵ��������£������ļ����Ǻε�С�����������Ӧ���ǣ�25��� ��10���꣬����1/4000��Ҳ����˵����һ�����������Ů��10����ij����У����ǿ��ܻ���������Ů��������ǧ��֮һ�Ļ��ᡣ��������10������û��������ֻ�е���һ��������һ��10���꣬����������һ����Ů��ʵ���ˡ� �Ӹ�����������Ҫ��4000��10���꣨����Լ4���꣩���ܳ��������ֵ�Ҫ��Ȼ����ʱ����ֻ��һ������ԳȺ�������200������ԳȺ������200�������Գ����ô4�������200���ͱ����200���ꡣҲ����˵�������200�����У�������200�������Գ�����п�����������һ�ԣ�3��Ⱦɫ�������ѧ�ǵ��Ϳ�ѧ���ޡ������ﻹ��һ�����������DZ���������һ��һŮ�����ҳɻ�������Ӻ�Ů������Ȼ������һ���ֹ۵Ĺ��ơ��ɴ����ǿ��Կ���ʵ����Щ�����Ƕ�ô����������������Ƕ�ô��С������ѧ�������������ġ�С�����¼������Ѿ��Dz����ܷ��������顣 ���˻�˵���Ͼ����Ǹ��ʵĹ��㣬��ʱҲ���������Ԥ��֮����漣�����ǣ����������������������ģ���������һЩ���ش�����⡣ ����ҳ |